Introduction
All organisms that we will investigate in the remaining animal labs have primary bilateral symmetry and are triploblastic, having three true germ layers: ectoderm, mesoderm and endoderm that form during gastrulation of the blastula stage of development. While radial symmetry may be well suited for sessile or slow-moving forms, animals that are active in seeking food, shelter and mates require a new body plan. Bilateral symmetry coupled with cephalization solves these problems. The anterior end moves forward and the posterior follows. The dorsal side is kept facing up and the ventral side is kept down and usually specialized for locomotion.
The bilateral grade of metazoans is further subdivided into two main divisions: the protostomes and deuterostomes, which are separated on the basis of a number of embryological differences. Evidence from sequence analysis of the small-subunit ribosomal gene suggests that some time after ancestral deuterostomes and protostomes diverged from one another during the Cambrian period, protostomes split into two “superphyla,” the Ecdysozoa and Lophotrochozoa. In this lab we will examine several lophotrochozoan phyla. First, we will take a close look at two of these phyla: Annelids and Mollusks. Then, we will do a more broad exploration of the diversity present in this group of organisms.
Annelida
The annelids generally have a well-developed coelom that acts as a hydrostatic skeleton: contraction of the muscular body wall against the incompressible coelomic fluid increases the internal pressure, creating stiffness and support. This type of skeleton also provides a means of locomotion; imagine a balloon filled with water (water is incompressible). If you squeeze the balloon at one end, the water must go somewhere, so it stretches the other end out, causing elongation.
Notice, however, that in this type of system there is little control over the movements: the single contraction causes a response along the whole length of the balloon (or worm). Because each segment of an annelid has its own set of circular muscles, and there is partial compartmentalization of the coelomic fluid by septa (infolded walls of tissue), the worm is able to control its movements. Annelids also possess longitudinal muscles at 90° angles to the circular musculature.
Directed elongation of the segments at the anterior end while simultaneously thickening (and anchoring) segments at the posterior end, followed by anchoring of the anterior segments, releasing the posterior segments, and then pulling the posterior end forward, allows the worm to move through the sediment. Phew, quite a long sentence! Try reading that again and imagining the sequence of events. Make a sketch that represents the sequence of events described. Think about when the circular and longitudinal muscles of each segment are in contracted vs. relaxed states, creating a wave-‐like sequence of contraction.
The members of this phylum, particularly the polychaete worms, have different types of life histories.
PHYLUM ANNELIDA
- Coelomate
- Bilateral symmetry
- Protostome development
- Metamerism (segmentation)
- Partial internal compartmentalization of the coelom by septa (except in leeches)
- Setae or chetae
- Closed circulatory system
- Excretion by metanephridia in most species
- Well-defined nervous system (brain and pair of ventral nerve cords)
- Hydrostatic skeleton
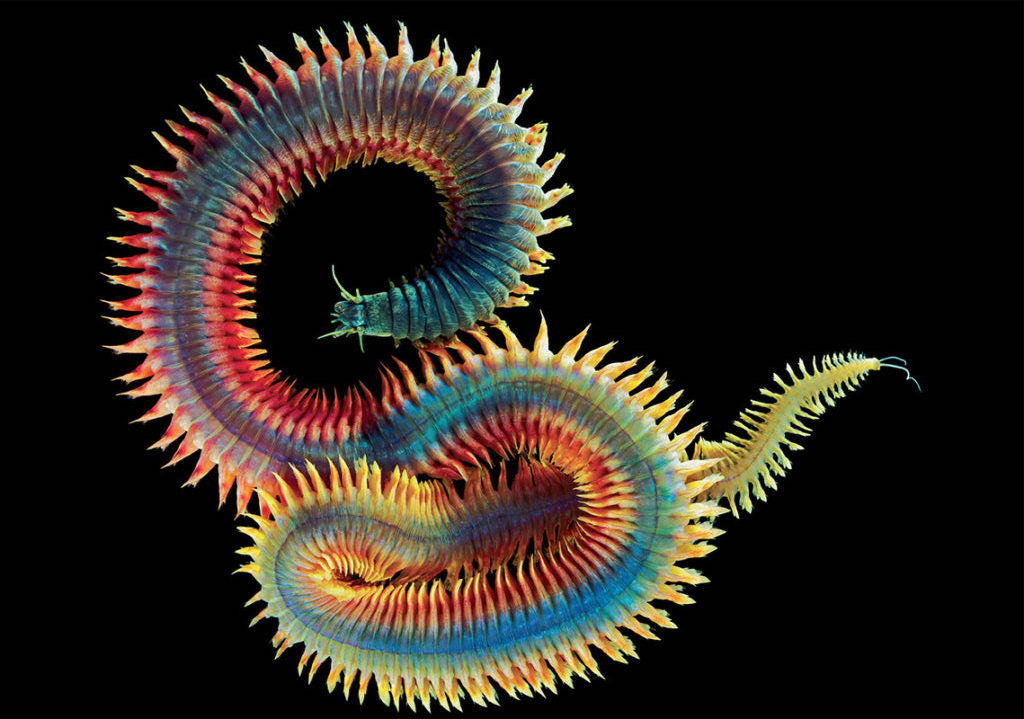
CLASS POLYCHAETA marine worms (>5000 spp.); free-‐living; parapodia (found on each side of most segments) and used for locomotion, respiration or feeding; setae located on the parapodia; dioecious (separate sexes); head often well-‐ developed; trochophore larva

CLASS OLIGOCHAETA includes earthworms (~3000 spp.); terrestrial or freshwater; no parapodia; short setae; monoecious (hermaphroditic); clitellum, a thickened area mid-‐body that secretes mucus and is used for sperm exchange; reduced head

CLASS HIRUDINEA leeches (~300 spp.); mainly freshwater, few marine and terrestrial in moist, warm areas; no parapodia or setae; hermaphroditic; clitellum present; suckers at anterior and posterior ends; ectoparasitic; carnivorous or detritivores (feeding on decaying matter); some leeches are free-‐living (not parasitic); coelom reduced—coelomic space virtually filled with connective tissue
Mollusca
The phylum Mollusca is the second most diverse phylum (after the arthropods) with 50,000 – 100,000 species including snails, slugs, clams, octopus and squid. Molluscs possess a coelom or fluid-‐ filled cavity between the body and the visceral organs. The molluscan coelom is relatively reduced, surrounding only the area around the heart. The sinuses of the open circulatory system of the majority of molluscs also serve as a hydrostatic skeleton used in locomotion by some species (especially burrowing bivalves). Others make use of the mantle (squid & octopus) for ‘jet propulsion’ or a muscular foot to generate pedal waves.
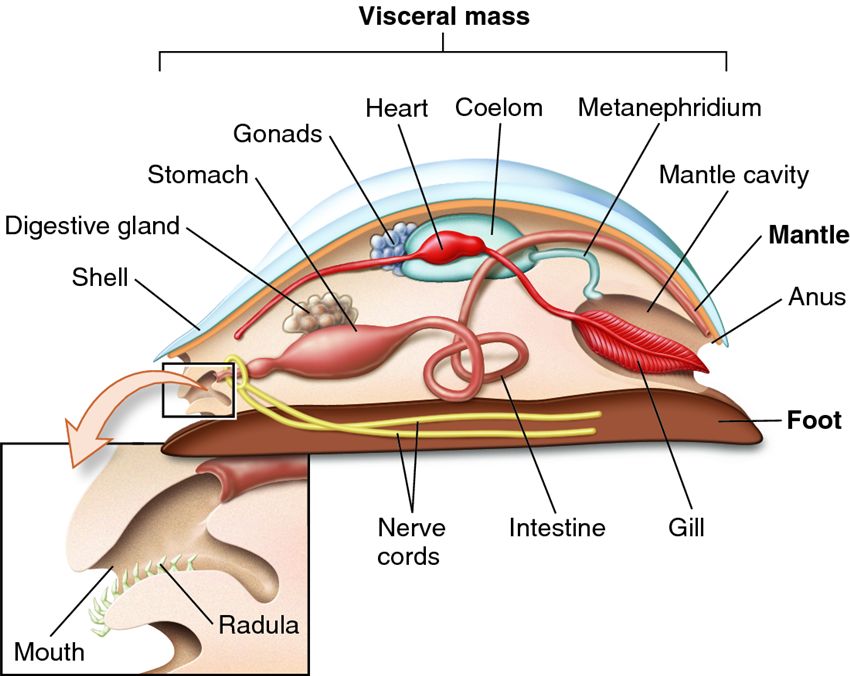
Another important characteristic of molluscs is the presence of a complete gut with a mouth at one end and an anus at the other. The one-‐way flow of material through the gut allows for regional specialization of the digestive tract. As the food moves along in one direction, certain portions of the digestive tract function in storage, mechanical grinding, and absorption. The unidirectional gut also allows the organism to keep eating while digesting and defecating. This more efficient mode of digestion is important, especially for large, very motile animals that require more energy. Animals that possess a complete gut also possess some type of circulatory system: when one region of the gut is specialized for nutrient absorption, the nutrients must be actively distributed to the rest of the body. As body size increases, simple diffusion is insufficient to deliver and remove respiratory gases, and a means of active respiration to transport oxygen to those tissues not in direct contact with oxygen becomes more important (i.e., a circulatory system).
PHYLUM MOLLUSCA
- Coelomate
- Bilateral symmetry
- Protostome development
- Body with three parts, including a muscular foot, a visceral mass, which contains the internal organs, and a mantle: a fold of tissue that generally secretes a shell
- Many possess a radula, a uniquely molluscan feature to scrape food
- Most molluscs pass through a trochophore larval stage, but there are additional larval forms also found in this phylum.
- Cilia are often present and are used in feeding & locomotion.
- Metanephridia are present in most species and function in osmoregulation and excretion.
- Molluscs are mostly dioecious (have separate males and females) and have an incredible diversity of body forms, habitats, and feeding habits.
- Seven major classes. We will examine four of these classes today:

CLASS GASTROPODA: snails, limpets, and slugs. Gastropods are a very diverse (>40,000 spp.) taxon. Generally well-‐developed head with eyes and tentacles; broad, creeping foot; coiled or cone-‐shaped shell, or no shell; body mass undergoes torsion during development. Gastropods can be marine, aquatic, or terrestrial.

CLASS BIVALVIA: clams, mussels, oysters, scallops. Two shell valves, right and left, secreted by large two-‐lobed mantle; Narrow foot used for burrowing; Head reduced or absent; No radula; Sense organs in mantle.

CLASS CEPHALOPODA: squid, octopus, nautilus, cuttlefish. Large well-‐developed head and nervous system with complex eyes and tentacles around the mouth derived from the foot; shell small or absent; active predators.

CLASS POLYPLACOPHORA: Marine animals (chitons) with oval body form and reduced head; Armor of 8 valve-‐shells over dorsal surface; radula present; Basic body plan of chiton probably represents the modern version of the “ancestral” Molluscan body plan.